|
HOME CONSERVATION C-TERMINAL ANALYSES DNA AND RNA SEQUENCES RESEARCH QUESTIONS RESULTS RESOURCES REFERENCES |
|
HOME CONSERVATION C-TERMINAL ANALYSES DNA AND RNA SEQUENCES RESEARCH QUESTIONS RESULTS RESOURCES REFERENCES |
|
|
|||||||||||||||||||||||||||||||||||||
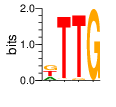
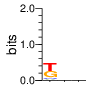
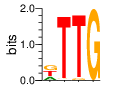
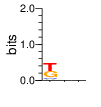
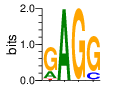
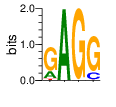
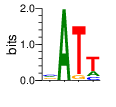
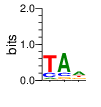
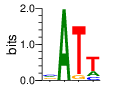
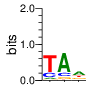
A first insight into the function of ZNF292 came from Lipkin (1993) where it was identified as an enhancer for the expression of growth hormone (GH) in rat pituitary. They compared the upstream GH sequences of several species and found a highly conserved sequence between two POU1F1 (Pit-1) binding sites. They named this sequence the Z-Box. Subsequent mutational analysis of the Z-box revealed its importance to efficient GH expression. Rat AGCACAAG-CTGTCAGTGG Bovine AGCACAGG-CTGCCAGTGG Porcine AGCACATGCCTGCCAGTGG Caprine AGCACAGG-CTGCCAGTGG Ovine AGCACAGG-CTGCCAGTGG GRch38 AGCACAAGCCCGTCAGTGG Consensus AGCACANG-CTGCCAGTGG Radiolabeled oligonucleotide was used to isolate from nuclear extracts a protein bound to the Z-Box which they named Zn-15 (human ZNF292). The sequence predicted 15 zinc fingers, but subsequent research determined that there are actually 16 VanderHeyden (2000). Here I will refer to zinc fingers by the 1-16 number scheme. Most cell lines tried showed Zn-15 binding activity suggesting it has functions outside of somatotrophs. Zn-15 has a predicted mass of 240Kd , but on SDS-PAGE appeared as 180 and 75Kd fragments using an anti-sera against C-terminal. They predicted that the 180Kd fragment was full-length and the 75Kd a proteolytic fragment. Anti-sera binding on multiple tissue sample showed ZN-15 to be widely expressed outside of pituitary in contrast to POU1F1 which is restricted to somatotrophs. Zn-15 was also found to bind to the T3RE of rat α-myosin heavy chain (human MYH6), but not other T3REs. It was able to compete for binding with T3/RXR heterodimers suggesting that Zn-15 may modulate some T3 receptor target genes. GH Z BOX AGCTGTCAGTGG αMHC T3RE AGGTGACAGGAG They found that a C-terminal fragment containing the last six zinc fingers bound to DNA whereas an N-terminal fragment containing the first seven zinc fingers did not. They mutated zf10-15 replacing the first histidine with a serine and found that zf10-12 were required for Z-box binding. Mutating zf13-15 had no effect on binding. Zn-15 Δ1-855 was found to efficiently stimulate transcription as did a larger deletion Zn-15 Δ1-1214 but not as effectively. Zn-15 Δ1215-2177 and a fragment containing only zinc fingers 10-12 (Zn-15 1214-1753) did not stimulate transcription. Together these results suggest a role for a portion of the protein extending from and including zf8 to but not including zf9 (855-1358) in cooperatively binding with another protein (likely POU1F1) to stimulate expression of GH. Protein locations are with respect to the recombinant protein used which may not be full length. Lipkin also showed that a mutated POU1F1 with the N-terminal removed (ΔN-POU1F1) as well as a point mutation involved in dwarfism (POU1F1 A158P) was unable to synergize with ZNF292. This work gives some insight into how the proteins may interact and allows us to ask whether or not ZNF292 may interact with other members of the POU1F1 family. Discussion Despite its apparent role in GH expression, ZNF292 is rarely referenced in the literature subsequent to Flynn (2006) if at all. For example, reviews of GH expression and associated pathologies fail to mention its role e.g. Mullins (2012). It seems that though it may be important to GH expression, the low required copy number (Wojtkiewicz, 2002) means haploinsufficiency is not an issue for a heterozygous loss of function mutation of ZNF292. Given its wide expression it is likely it has a different role outside of somatotrophs where heterozygous LGD mutations are pathogenic. We also need to consider the complex metabolic role growth hormone plays in the human body. It is tightly regulated with basal levels achieved during the day along with spikes, and highest levels being reached during REM sleep (Mullin, PE 2012). If ZNF292 is involved in the fine control of GH expression it could have affects more subtle than simply a slowed over-all growth profile. Mutations that affect development of the pituitary (hypopituitarism) have been associated with midline malformations such as holoprosencephaly, septo-optic dysplasia and Kallmann Syndrome. Symptoms of KS include high arched palate, cleft lip/palate, dental agenesis, sensorineural hearing loss and hyposmia or anosmia. Growth hormone deficiency has been correlated with variations in brain structures including the corpus callosum, left globus pallidus, thalamus and hippocampus that lead to deficits in cognition and motor function (Webb, 2018). GH deficiency has been linked to lower levels of brain N-acetylaspartate (van Dam, 2005). Low NAA in the brain has been observed in children with ASD and intellectual disability (Corrigan 2013). A Deeper Look at the ZRE-box The exact 19bp of the human ZRE box as described by Lipkin (1993) appears in several other locations in the growth hormone locus on chromosome 17. POU1F1 binding sites may also be present in some of these promoters (Jacobsen, 1997) Locations of Human ZRE box (AGCACAAGCCCGTCAGTGG) found in GRCh38p10 Human ZRE-box Possible POU1F1 binding site TAAAT or (A/T)(A/T)TATNCAT (Jacobsen, 1997) GH1 (somatotropin isoform 1 precursor) 154 bp at 5' side AACACTGGTGACGGTGGGAAGGGAAAGATGACAAGCCAGGGGGCATGATCCCAGCATGTGTGGGAGGAGC TTCTAAATTATCCATTAGCACAAGCCCGTCAGTGGCCCCATGCATAAATGTACACAGAAACAGGTGGGGT CAACAGTGGGAGAGAAGGGGCCAGGGTATAAAAAGGGCCCACAAGAGACCAGCTCAAGGATCCCAAGGCC CAACTCCCCGAACCACTCAGGGTCCTGTGGACAGCTCACCTAGCTGCAATG GH2 (growth hormone variant isoform 2 precursor) 154 bp at 5' side TGCACAACCCTCACAACGCTGGTGATGGTGGGAAGGGAAAGATGACAAGTCAGGGGGCATGATCCCAGCA TGTGTGGGAGGAGCTTTATCCACTAAATTTAGCACAAGCCCGTCAGTGGCCCCAGGCCTAAACATGCAGA GAAACAGGTGAGGAGAAGCAGCGAGAGAGAAGGGGCCAGGTATAAAAAGGGCCCACAAGAGACCAGCTCA AGGATCCCAAGGCCCAACTCCCCGAACCACTCAGGGTCCTGTGGACAGCTCACCTAGCGGCAATG CSH1 (chorionic somatomammotropin hormone 1 precursor) 155 bp at 5' side TGATGGTGAGAAGGGAAAGACGACAAGCCAGGGGGCATGATCCCAGCATGTGTGGGAGGAGCTTCTAAAT TATCCACTAGCACAAGCCCGTCAGTGGCCCCATGCATAAATGTACACAGAAACAGGTGGGGTCAAGCAGG GAGAGAGAACTGGCCAGGGTATAAAAAGGGCCCACAAGAGACCGGCTCTAGGATCCCAAGGCCCAACTCC CCGAACCACTCAGGGTCCTGTGGACAGCTCACCTAGTGGCAATG CSH2 (chorionic somatomammotropin hormone 2 isoform 2 precursor) 155 bp at 5' side ACCCTTAAAGAGAGGACAAGTTGGGTGGAGTCTGTGGCTGACACTCTGTGCACAATCCTTACAACACTGG TGATGGTGAGAAGGGAAAGACGACAAGCCAGGGGGCATGATCCCAGCATGTGTGGGAGGAGCTTCCAAAT TATCCATTAGCACAAGCCCGTCAGTGGCCCCATGCATAAATGTACACAGAAACAGGTGGGGTCAAGCAGG GAGAGAGAACTGGCCAGGGTATAAAAAGGGCCCACAAGAGACCGGCTCTAGGATCCCAAGGCCCAACTCC CCGAACCACTCAGGGTCCTGTGGACAGCTCACCTAGCGGCAATG CSHL1 (chorionic somatomammotropin hormone-like 1 isoform X1) 154 bp at 5' side TAAAGAGAGGACAAGTTGGGTGGTGTCTGTGGTTGACACTCTGTGCACAACCCTCACAACGCTGGTGACG GTGGGAAGGGAAAGATGACAAGCCAGGGGACATGATGCCAGCATGTGTGGGAGGAGCTTCCAAATTATCC ATTAGCACAAGCCCGTCAGTGGCCCCATGCATAAATGTGCACAGAAACAGGTGGGGGCAACAGCGAGAGA GAAGGGGCCAGGGTATAAAAAGGGCCCACAAGAGACCAGCTCCAGCATCCCAAGGCCCGACTCCCCGCAC CACTCAGGGTCCTGTGGACAGCTCACCTAGCGGCAATG Playing with Fingers PWMs calculated using zf.princeton.edu with Expanded Linear SVM or Random Forest Regression based on B1H dataset zf10 1904 1927 GRCh38 canonical FVCQNQGCNYSAMTKDALFKHYGKIH (human) FVCQNQGCNYSAMTKDALFKHYGKIH (rat) ************************** FICQETDCTYCAMTKDALFKHYAKVH (xenopus) *:**: .*.*.***********.*:* 1 gnomAD v2.1 Controls Mutation N PolyPhen Grantham Phast GERP CADD p.Phe1920Ile 1 0.976 21 0.933 6.17 16.61 Human
ZF10 AGCACAAGCCCGTCAGTGG TCGTGTTCGGGCAGTCACC ZF11 .. ... AGCACAAGCCCGTCAGTGG TCGTGTTCGGGCAGTCACC ... ZF12 AGCACAAGCCCGTCAGTGG TCGTGTTCGGGCAGTCACC |